 m.quanpro.cn
m.quanpro.cn
媒体:国家动物博物馆 作者:国家动物博物馆
专业号:野生动物保护协会
2021/11/22 14:26:41
首页 > >
两岸猿声今何在——在我国艰难生存的长臂猿
长臂猿是一类分布于东南亚地区的无尾灵长类动物,它们与猩猩类并称为类人猿。人们熟悉长臂猿多是由于李白的诗句“两岸猿声啼不住,轻舟已过万重山”。这个不朽的诗句说明,在李白生活的唐代,长臂猿的分布区已经出现在长江两岸。如今,长臂猿在我国的分布区已大幅退缩,仅存于云南、广西和海南。

长臂猿生存的热带雨林环境 孙忻/摄
长臂猿以长臂无尾、林间荡行、一夫一妻、雌雄异色、鸣声嘹亮而著称。长臂猿的上肢明显长于下肢,且肩关节和腕关节均为及其灵活的球窝关节,这种关节结构可以让长臂猿以独特的大幅荡行方式极快的游走于林冠上层,如风似电。因此在我国云南地区的老百姓把长臂猿称为“风猴”,可见其移动速度之快。
长臂猿为典型的一夫一妻制物种,近年的研究表明,在我国生存的冠长臂猿属(西黑冠长臂猿、东黑冠长臂猿、海南长臂猿)实行着稳定的一夫二妻婚配制度。更为独特的是,长臂猿是少有的存在“性二型”特征的哺乳动物,大多数长臂猿的雄性毛色为黑色,而雌性多为米黄色。同一种长臂猿其毛色也会有年龄性别的差异,以西黑冠长臂猿为例,新生幼猿(1周龄)雌雄两性都为淡黄色,随着年龄的增长(1 月龄)逐渐变成金黄色,在1岁左右雌雄都从黄色逐渐变成黑色;雌性个体在性成熟时身体的大部分再次变成黄色,仅在腹部和头顶保留部分黑毛,而雄性个体成年后仍然保持黑色。
鸣叫是长臂猿的典型行为,长臂猿都有清晨鸣叫的习性,鸣声高亢嘹亮,在几公里以外都可以听到。长臂猿鸣声的主要功能为群间的隔离、群体的定位、资源防御(如领域、食物资源或配偶)、群体的形成与维持、配偶吸引、配偶稳固和宣传配偶信息等。长臂猿的鸣叫一般由雄性开始,之后雌性加入。雌性发出固定刻板而且频率非常高的叫声,称之为“激动鸣叫”,与雄性配合组成二重唱。除克氏长臂猿和穆氏长臂猿外,群体中的成年雌雄个体可以配合发出独特的二重唱。这种二重唱的功能之一就是维持配对之间的关系,并向同种的其它个体传播有关配对的稳定性的信息。研究发现,新组建的家庭必须在尽量短的时间内发出配合默契的二重唱。因为不完美的二重唱可能反映了配对的不稳定,进而引起同性个体对配偶的竞争。
长臂猿多生活在植被保存完好的原始常绿阔叶林和热带雨林中,是典型的热带雨林指示物种和旗舰物种,长臂猿在某地的消失意味着该地区生态系统受到了较为严重的破坏。长臂猿以果实为主要食物,辅以嫩叶、花、茎等植物性食物和极少量的昆虫、鸟卵、幼鸟、蜥蜴等动物性食物。在我国云南无量山,科研人员还发现西黑冠长臂猿捕食大鼯鼠。在一项长达14个月累计845小时的野外研究中,观察发现西黑冠长臂猿捕捉大鼯鼠11次,成功4次,被捕食的4只大鼯鼠均为幼体和压成体。
植物果实在长臂猿的食谱中占据了77%以上的比例,表明这类灵长类动物对于果实的倚赖性较强,受到破坏后的次生林往往无法满足长臂猿对食物的需要。这种食物结构也限制了长臂猿种群的恢复与增长。毕竟,果实在热带雨林中的分布存在明显的季节性和稀缺性。
随着分子生物学方法的介入,结合形态、行为、鸣声等特征的大量研究,长臂猿科现分为冠长臂猿属、长臂猿属、白眉长臂猿属和合趾猿属等4属,共20种,分布在我国的有6种,分别为西黑冠长臂猿、东黑冠长臂猿、海南长臂猿、北白颊长臂猿、白掌长臂猿和高黎贡白眉长臂猿,而西白眉长臂猿在我国的分布还属存疑状态。
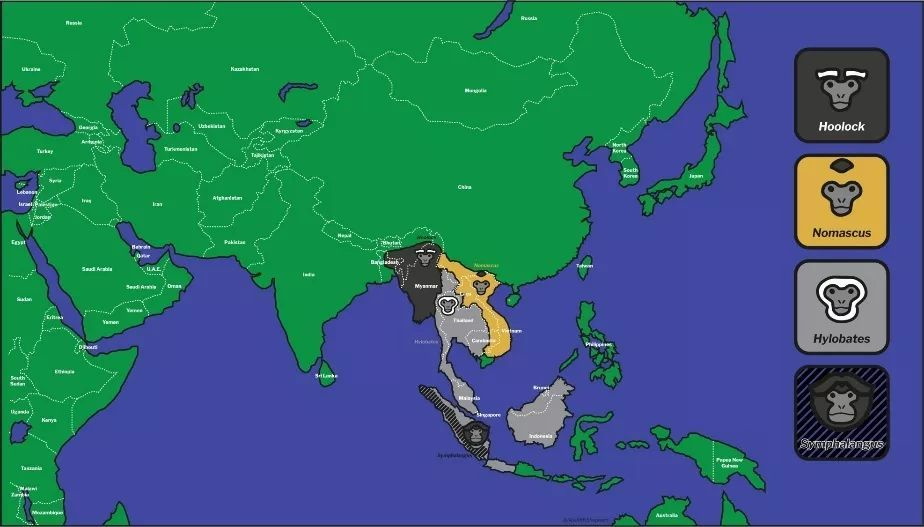
Hoolock白眉长臂猿属、Nomascus冠长臂猿属、Hyiobates长臂猿属、Symphalangus合趾猿属分布图
图片来自IUCN长臂猿专家组
本文将分别讲述在我国分布的6种长臂猿的生存状况。
冠长臂猿属
冠长臂猿属物种的突出特征是雄性头顶有短而直立的冠状簇毛,包括西黑冠长臂猿、东黑冠长臂猿、海南长臂猿、北白颊长臂猿、南白颊长臂猿、北黄颊长臂猿和南黄颊长臂猿,共7种。冠长臂猿属物种的分类经历了较长时间的研究,目前7种可分为无白颊型和白颊型。无白颊型分为西黑冠长臂猿、东黑冠长臂猿、海南长臂猿等3种,白颊型分为北白颊长臂猿、南白颊长臂猿、北黄颊长臂猿和南黄颊长臂猿等4种,最后从亚种提升为物种的是北黄颊长臂猿,时间是2010年。
非常有趣的是,我国科研人员发现3种无白颊型冠长臂猿都可以形成稳定的“一夫二妻制”社会结构,这一点与其它长臂猿“一夫一妻制”社会结构明显不同,其产生的原因尚不明确。至于4种白颊型冠长臂猿的社会结构是否也是“一夫二妻制”还是个谜。这些长臂猿分布在中南半岛东部的越南、老挝、柬埔寨等地,科研机构还未对这4种白颊型冠长臂猿开展较为深入的研究。
1、西黑冠长臂猿
西黑冠长臂猿分布于我国云南中部到南部、越南北部和老挝西北部。在我国,云南中部的哀牢山区和无量山区是西黑冠长臂猿的主要分布区。2012年的文献显示,目前全球西黑冠长臂猿的种群数量约为300群,其中滇中哀牢山约170群,无量山有87群;滇南金平芭蕉河有2群、西隆山可能有1-2群,绿春黄连山可能有1-3群;滇西永德大雪山有4群;越南北部的穆庆寨(Mu Cang Chai)和山萝(Son La)分别不少于14群和6群;老挝北部的南哈地区(Nam Ha)不少于1群,南坎(Nam Kan)地区至少13群。中国生存着90%以上的现生种群,种群数量约1300只,因此在西黑冠长臂猿的保护中起着至关重要的作用。

西黑冠长臂猿雄性 摄影/赵超
我国科研人员对西黑冠长臂猿的监测工作开始于2003年。初期以鸣声监测为主,经过1 年多的鸣声监测,研究人员初步掌握了长臂猿群体的领域范围和群体组成。2005年,中国科学院昆明动物研究所蒋学龙研究员的博士生范朋飞在景东无量山成功习惯化我国第一个长臂猿群体G3 群,对深入研究该物种具有重要的奠基意义。
2003-2005年,范朋飞等对无量山大寨子西黑冠长臂猿群体的结构和组成进行了观察。当一个群体在早晨鸣叫或依次通过树冠时, 记录群体的结构和组成。研究发现,每个群体都由1个成年雄性和2个成年雌性及其后代组成。这一点和我们熟知的长臂猿“一夫一妻”制明显不同。在3个群体中,西黑冠长臂猿中都有两个成年雌性成功繁殖了后代,且同一群体内两个成年雌性间无攻击或等级行为。至此,西黑冠长臂猿稳定的“一夫二妻” 制社会结构浮出水面。

西黑长臂猿冠雌性 赵超/摄
2010 年,中国科学院昆明动物研究所蒋学龙研究员的博士生管振华成功习惯化G3群的相邻群体G2群,为群体间对比研究提供条件。通过分析理毛行为和空间近距离,科研人员构建了G2群和G3群的社会网络结构,发现2个雌性主动相互理毛以维持其社会关系,深入探讨了长臂猿社会关系的维持机制,对研究长臂猿“一夫二妻制”社会结构的形成具有重要启示。2012年无量山国家级自然保护区、哀牢山国家级自然保护区景东管理局的工作人员成功习惯化第三个群体G4群。至此,大寨子长臂猿小种群3个群体全部被成功习惯化。10多年的野外监测为研究者对该种群的基础研究提供了丰富且重要的研究数据。
尽管西黑冠长臂猿是在我国分布的6种长臂猿中种群数量最多的一种,但是仍面对着栖息地大幅退缩和破碎化的迥境。2020年的文献指出,相比五十年代,西黑冠长臂猿当前的潜在分布区已经退缩69%,且大生境斑块距离种群潜在分布区中心越来越远,各个种群的海拔分布上限并没有显著提高,且海拔下限均有不同程度的提升。这说明,人类影响是驱动该物种分布区退缩的重要因素。
2、东黑冠长臂猿
东黑冠长臂猿历史上曾分布于红河以东的中国南部和越南北部,自上个世纪50年代起一度被认为已经从中国灭绝,60年代后越南也没有该物种分布的确切消息。直到2002年,野生动植物保护国际(Fauna & Flora International, FFI)在越南北部高平省Trung Khanh(重庆县)的喀斯特森林中重新发现东黑冠长臂猿。2006年5月,广西大学周放教授在我国广西靖西市邦亮林区录到了东黑冠长臂猿的叫声。2006年9月,香港嘉道理农场和靖西县林业局组织的调查队于在中国广西靖西县与Trung Khanh(重庆县)交界的森林中发现了3个东黑冠长臂猿群体。随后,中越跨边境保护工作开启。当地政府针行动迅速,对邦亮林区采取了紧急保护措施,包括消除烧炭、开荒等人为活动。

东黑冠长臂猿雌性与幼猿 摄影/赵超
2007年9月,中越两国开展联合调查,共发现18群约110只。分布于中国的东黑冠长臂猿有4 群约23只,其中3群长臂猿跨中越边境生活。2008年5月越南在林区修建国界巡护路对其中一群长臂猿造成了严重影响,该群体的活动范围发生了明显改变,进而导致另一个群体的活动范围完全退入越南境内。
我国科研机构对东黑冠长臂猿的监测工作始于2007年,由于地形条件限制,无法成功被习惯化,但喀斯特地区开阔的视野条件和针叶林为研究者提供了有利的观察条件。初期通过鸣声监测获取录音文件进行声谱分析,从鸣叫特征上支持了东黑冠长臂猿、西黑冠长臂猿和海南长臂猿的分类地位; 随着监测工作的持续开展,研究者进一步研究了该种群的群体组成、活动时间分配以及栖息地特征,结合物候监测分析了该种群对贫瘠的喀斯特地区森林的适应。
2009年7月,广西壮族自治区人民政府批复建立广西邦亮东黑冠长臂猿自治区级自然保护区,保护区总面积6530公顷。2013年12月25日通过国务院办公厅批复正式晋升为国家级自然保护区。保护区成立以来,就以监测巡护、社区参与以及与越南保护区建立联合保护等多种方式加强对东黑冠长臂猿及喀斯特季雨林的保护。

东黑冠长臂猿雄性 赵超/摄
2015年1月,科研人员发现在中国境内形成了由1只成年雄性、2只成年雌性和1只婴猿组成的新群体(GL 群)。这是自2006年该物种在中国被重新发现后,首次在中国境内发现形成的新群体,也是唯一一群完全生活于我国的东黑冠长臂猿家庭。研究表明,东黑冠长臂猿也存在稳定的“一夫二妻制”家庭结构。虽然GL 群是一个新形成的群体,但这个家庭也包含了两只成年雌性。
2016年9月中越两国再次开展联合调查,调查结果为21-22群129-136只,中国境内4群26只。由于该物种目前仅存一个种群,IUCN 将其列为极度濒危(Critically Endangered)物种,世界上最濒危的25 种灵长类之一。
时至今日,在我国出现的东黑冠长臂猿共5群33只,其中的1群完全生存在我国境内,其余4群则在中越国境线上穿梭。由“缤纷自然”拍摄的纪录片《方舟.东黑冠长臂猿》是世界上首部关于东黑冠长臂猿的纪录片,该片获得中国野生生物视频年赛大奖,影片把世人的眼球聚焦到这种濒危的野生动物身上。中山大学范朋飞教授的科研团队在广西邦亮保护区卓有成效的科研工作、民间组织“云山保护”对东黑冠长臂猿的宣教和调查工作,以及当地保护区与越南高平重庆自然保护区的联合保护工作令东黑冠长臂猿的未来充满希望。
3、海南黑冠长臂猿
海南黑冠长臂猿是我国唯一一种特有的长臂猿,仅分布于我国海南霸王岭保护区,目前种群数量为5群35只,是全球最为濒危的灵长类。2020年,一篇题为The Genetic Status of the Critically Endangered Hainan Gibbon (Nomascus hainanus): A Species Moving Toward Extinction的论文表明,海南黑冠长臂猿的遗传多样性极低,这种最濒危的灵长类动物正在经历着高水平的近亲繁殖,加之面对着高达99.9%的栖息地丧失和破碎化,以及新生猿宝宝极不平衡的性比问题,海南黑冠长臂猿面临着极高的灭绝风险。世界自然保护联盟(IUCN)2007年宣称,海南黑冠长臂猿是21世纪最有可能灭绝的灵长类。

海南黑冠长臂猿雄性 摄影/赵超
在上个世纪50年代,海南黑冠长臂猿仍然分布在海南岛的12个县,种群数量超过2000只。但是到了1978年,海南黑冠长臂猿的数量已经下降到了30-40只,仅分布于霸王岭和黎母岭。2002年9月,贵州师范大学周江团队开始对霸王岭的海南黑冠长臂猿种群进行监测研究,此时该种群为2群13只。2007年底,其种群数量增长到16-20只,但是依然只有2个家庭群。这两个群体都是“一夫二妻”的社会结构,但其中一只成年雌性由于年龄过大可能已经失去繁殖能力,这意味着海南黑冠长臂猿这一物种仅剩下3只具有繁殖能力的雌性。研究发现,海南黑冠长臂猿2个群体的领域面积分别达到9. 87 km2和5. 48 km2,远大于其它种类长臂猿的领域面积,分析表明栖息地质量不高及食物资源差是造成海南黑冠长臂猿需要大面积领域的主要原因。
早期的文献表明,从1986 年开始,霸王岭2个家庭群都有2只成年雌性。1994 年保护区内新增一个家庭群,为一夫一妻制。到了1997 年,霸王岭保护区内的3 群长臂猿均为一夫二妻的社会结构。2002年开始,贵州师范大学周江团队在霸王岭国家级自然保护区作研究时,发现的当时仅有的2个家庭群均为“一夫二妻”的社会结构,而于2011年新形成的一个家庭群也是一夫二妻。因此,海南黑冠长臂猿和其它两种黑冠长臂猿一样,存在稳定的“一夫二妻制”社会结构。

海南黑冠长臂猿母与子 赵超/摄
低质量的栖息地、较长的生殖间隔及较低的遗传多样性是制约海南黑冠长臂猿种群生存和发展的主要原因。2011年6月,虽然个体数量未有明显增长,但群体已经扩大为3群。2013年,种群数量增长为3群20只和6只独猿。2015年6月,第4群形成,种群数量为4群25只。2020年,种群数量增长为5群33只。
2021年3月,随着2只小猿的出生,种群数量达到了5群35只。第5个家庭群出现在距离原栖息地昌江黎族自治县霸王岭林区10公里的东崩岭,这证明海南黑冠长臂猿的栖息地面积在扩大,且新栖息地自然条件能够支持其生存。
从2002年的2群13只到2021年的5群35只,海南黑冠长臂猿保护初见成效,但这一数量仍远低于生态学上种群能够持续生存的最小种群数,种群复壮依然艰巨。为保护这一濒危物种脱离灭绝困境,过去几十年间,海南省霸王岭林业局和海南霸王岭自然保护区联合霸王岭森林公安实施了最为严格的栖息地保护措施。2005年,海南省霸王岭林业局和嘉道理农场暨植物园共同组建长臂猿监测队,发展至今共有19位队员,负责长臂猿监测任务,同时统计家庭群数、记录个体年龄、拍摄图像资料、采集新鲜猿粪,为海南黑冠长臂猿的科学研究提供了第一手材料。在栖息地修复方面,海南省林业部门自2005年起改造修复长臂猿栖息地5000多亩,种植超过30万株长臂猿喜食的乡土树种;2004年至2012年,嘉道理农场暨植物园在南叉河种植2300亩,53种8万多株长臂猿喜食的南酸枣、秋枫、榕树等树种,并每年抚育,如今已有成效。在山体滑坡处种下成片本土树苗和长臂猿喜食树种,并为长臂猿搭建生态绳桥。目前树种的高度已足够长臂猿活动,绳桥等生态廊道达到七条,并被长臂猿频繁使用。
2021年8月,“海南长臂猿保护国际研讨会”召开,并发布《海南长臂猿保护行动计划纲要》。与会专家一致认为,在加强保护力度,没有重大自然灾害的前提下,以国内科研力量为主体,整合全球力量开展联合攻关,海南黑冠长臂猿的数量有望在15年左右实现翻番并持续增长。2021年10月12日,国家主席习近平以视频方式出席《生物多样性公约》第十五次缔约方大会领导人峰会并发表主旨讲话。在讲话中,习近平宣布,中国正式设立三江源、大熊猫、东北虎豹、海南热带雨林、武夷山等第一批国家公园。海南热带雨林国家公园的建立,将为海南黑冠长臂猿这一极度濒危的物种带来新的机遇。
4、北白颊长臂猿
北白颊长臂猿是国家一级保护动物,IUCN 将其列为极度濒危 ( Critically Endangered) 物种。除中国外,该物种还分布于老挝和越南北部,但这两个国家的种群数量不详。北白颊长臂猿曾经广泛分布于西双版纳的热带雨林和沟谷雨林中。1958年,在勐腊县城每天早晨都能听到不同方位传来的长臂猿叫声。20 世纪 60 年代,中国曾分布有大约1000 只北白颊长臂猿,但是在80年代其种群数量迅速下降到 100 只左右。随着西双版纳热带雨林的大规模开发和橡胶的大量种植,北白颊长臂猿种群数量迅速下降,至1989年西双版纳仅生存在七个地区,每个地区仅存 1 -2 群,种群数量不足40只。

北白颊长臂猿雄性 赵超/摄
随着西双版纳保护区的成立,西双版纳残存的热带雨林得到了有效的保护,但受到偷猎和小种群效应的影响,这些局部种群先后灭绝。2008年12月范朋飞团队在西双版纳进行调查时,发现北白颊长臂猿可能仅在三个相互隔离的区域中,数量不足10只。2011年12月,范朋飞团队在西双版纳的再次调查显示,北白颊长臂猿可能已经在我国灭绝。

北白颊长臂猿一家 引自云山保护
虽然我们现在可以在西双版纳野象谷看到北白颊长臂猿,但那是野象谷景区人工饲养的几只,并非野外种群。即便如此,这几只人工饲养的北白颊长臂猿可能成为该物种在中国的野化和重新引入保留了最后的一丝希望。
长臂猿属
长臂猿属是20种长臂猿科动物中最为兴旺的家族,包括戴帽长臂猿、白掌长臂猿、敏长臂猿(白手长臂猿)、穆氏长臂猿(灰长臂猿)、阿氏灰长臂猿、白须长臂猿、北婆罗洲灰长臂猿、银白长臂猿(爪哇长臂猿)和克氏长臂猿,共9种。长臂猿属物种的分布范围最广,包括中南半岛中部到西南部、马来半岛、苏门答腊岛、婆罗洲和爪哇岛。
与白眉长臂猿属和冠长臂猿属不同,长臂猿属中的很多物种的雌雄毛色一致,没有区别。白掌长臂猿雌雄更是具同时有黑色和米黄色毛色,可谓“安能辨我是雌雄?”
5、白掌长臂猿
白掌长臂猿曾经分布于我国云南的沧源、西盟和孟连等县,为该物种分布的北限。中国科学院昆明动物研究所马世来和王应祥对采自孟连的 4 个标本和白掌长臂猿其它 4 个亚种进行了比较,根据孟连标本背毛长、阴毛和体色与其它亚种有明显差异等特征将我国的白掌长臂猿命名为一新亚种——云南亚种。

白掌长臂猿浅色型 孙忻/摄
1985年,白掌长臂猿在我国的数量仅为5群19-27只。南滚河保护区被认为是我国白掌长臂猿最后的栖息地,到了上世纪末,保护区内的长臂猿种群数量已经下降至3群10只左右。2007 年 11 月,瑞士苏黎士大学和中国科学院昆明动物研究所组织的联合考察队,对南滚河国家级自然保护区进行了为期两周的野外调查。此次调查没有听到白掌长臂猿的叫声,且根据访问调查的结果,2000 年以后再也没有人听到过白掌长臂猿的叫声。

白掌长臂猿深色型 赵超/摄
虽然白掌长臂猿在我国已经灭绝,但是白掌长臂猿在泰国的生存状况良好,在岗卡章、考亚等保护地都生存着数量不少的白掌长臂猿。2015年,我两次前往泰国观赏野生动物,同时拜访了泰国国立朱拉隆功大学生物系主任、博士生导师、泰国国家灵长类研究中心主任苏琴达.马来维基诺得(Suchinda Malaivijitnond)教授,也拜访了她的博士生日本京都大学的松平一成(Kazunari Matsudaira),松平博士一直在考亚国家公园研究白掌长臂猿和戴帽长臂猿。也是在那个时候,我才知道白掌长臂猿的雌雄两性都有淡色型和深色型的毛色,也就是说,我们不能从成体的黑色和米黄色毛色来区分它们的性别。更加令我惊奇的是,考亚国家公园里生活着800只白掌长臂猿,而白掌长臂猿在我国却已经绝迹多年了!
白眉长臂猿属
白眉长臂猿属物种的突出特征是面目有明显的白眉,包括西白眉长臂猿、东白眉长臂猿和高黎贡白眉长臂猿(天行长臂猿),共3种。白眉长臂猿曾被认为只有一个物种,分布于缅甸、印度、孟加拉和我国,东部白眉长臂猿和西部白眉长臂猿之前被认为是白眉长臂猿的两个亚种。2006年,IUCN灵长类专家组会议根据东部白眉长臂猿和西部白眉长臂猿特征的显著不同,将东白眉长臂猿和西白眉长臂猿提升为两个独立物种。
2017年,中山大学范朋飞教授团队科学命名了一种分布于我国云南地区的白眉长臂猿——高黎贡白眉长臂猿,也称天行长臂猿,这一发现将白眉长臂猿属的物种数上升为3种。在我国藏南地区有可能分布有白眉长臂猿的一个种群,曾经被认为是东白眉长臂猿,但是最新的研究表明其为西白眉长臂猿。由于西白眉长臂猿在我国的分布缺乏确凿的证据,因此未列入在我国分布的长臂猿名录中。
6、高黎贡白眉长臂猿(天行长臂猿)
生活在我国云南的白眉长臂猿原先被认为是东白眉长臂猿。东白眉长臂猿长臂猿分布于缅甸东部、印度东北部和我国云南西部高黎贡山地区,其中缅甸数量为30000-37000只(2013年数据)、印度数量为719只(2010年数据)、我国数量约为150只。中山大学范朋飞教授的科研团队长期开展我国长臂猿的研究工作,发现分布在我国云南的东白眉长臂猿与分布在缅甸和印度的“兄弟”在外形、叫声等方面有所不同,在进行了DNA测序工作之后,结果显示生活在我国云南的白眉长臂猿与生活在缅甸的东白眉长臂猿约在50万年之前就开始独立演化。范朋飞教授与美国、英国、德国和澳大利亚五国科学家组成的研究团队耗费近10年的时间,确认分布于云南高黎贡山的白眉长臂猿是一个新种——高黎贡白眉长臂猿(天行长臂猿),并将这些成果投稿到American Journal of Primatology,于美国东部时间2017年1月11日在线发表,标志着学术界认可长臂猿家族新成员的“诞生”。高黎贡白眉长臂猿(天行长臂猿)的科学发现,将长臂猿的物种数上升到了20种。

高黎贡白眉长臂猿(天行长臂猿)雌性 孙忻/摄
高黎贡白眉长臂猿是白眉长臂猿属的第3个物种,主要分布于怒江与伊洛瓦底江之间的中缅交界地区,即东部以怒江为界,西部以伊洛瓦底江及其上游恩梅开江为界。国内仅分布于怒江以西的高黎贡山南段保山市隆阳区、腾冲县和德宏州盈江县。和很多新发现的物种一样,高黎贡白眉长臂猿刚被命名就成为了濒危物种。高黎贡白眉长臂猿种群数量仅150只左右,被分割在若干破碎化的栖息地中。
我国高黎贡白眉长臂猿的监测先后在云南高黎贡山赧亢片区和大塘东部开展,2007-2008 年西南林业大学的研究团队对赧亢片区 1个群体和 1只雌性独猿进行了半年短期跟踪监测。2010年,范朋飞团队继续跟踪监测了这个群体并研究了时间分配及其领域行为。2009年10月,研究人员对高黎贡山大塘保护站辖区的3个群体进行鸣声监测,分析了该地区长臂猿的鸣叫特征。2012 年分布于板厂片区3个家庭群中的1群被成功习惯化,并被持续跟踪监测至今。

高黎贡白眉长臂猿(天行长臂猿)雄性 孙忻/摄
栖息地之间的阻隔导致的栖息地破碎化已经严重影响到长臂猿的生存。以生活在高黎贡国家级自然保护区赧亢站的高黎贡白眉长臂猿为例,该区域总共5只,包括1个家庭群4只和1只雌性孤猿。雌性孤猿已满20岁了,20岁对于人类而言是花样年华,而对于一只长臂猿来说则是迟暮之年。之所以成为孤猿是因为没有成年雄性与她配对,即便家庭群中的雄性小猿在成年后与孤猿组建家庭,孤猿也由于年龄问题很难生育后代(长臂猿寿命在30岁左右,10岁成年,才可以参与繁殖)。更加遗憾的是,赧亢站唯一一个家庭群中的“老大”被林雕捕食,“老二”曾被认为是一只雄性,所有的目光都希望“老二”成年后可以和那只“孤猿”组成新的家庭,哪怕“孤猿”不能生育。然而,在2019年,“老二”那身黑色的皮毛蜕变成为黄褐色,“老二”是一只雌性。我们可以预见,随着这几只长臂猿的逐一老去,赧亢站的高黎贡白眉长臂猿终将走向灭绝。其它地区的高黎贡白眉长臂猿的境遇也大致如此。此外,非法偷猎、栖息地丧失和退化(包括毁林开荒、伐木、放牧、林下种植草果等)也是导致我国东白眉长臂猿致危的重要因素。
致敬科研团队
在我国,长臂猿科所有物种均是国家Ⅰ级重点保护野生动物。全世界共有20种长臂猿,我国生存有6种长臂猿,三是其中2种约在20年前灭绝。目前生存在我国的4种长臂猿中,海南黑冠长臂猿、东黑冠长臂猿的种群数量都在40只以下,这个数量还不到小学一个班学生的数量。4种长臂猿的总数还不足1500只,濒危程度远超人们熟知的大熊猫。
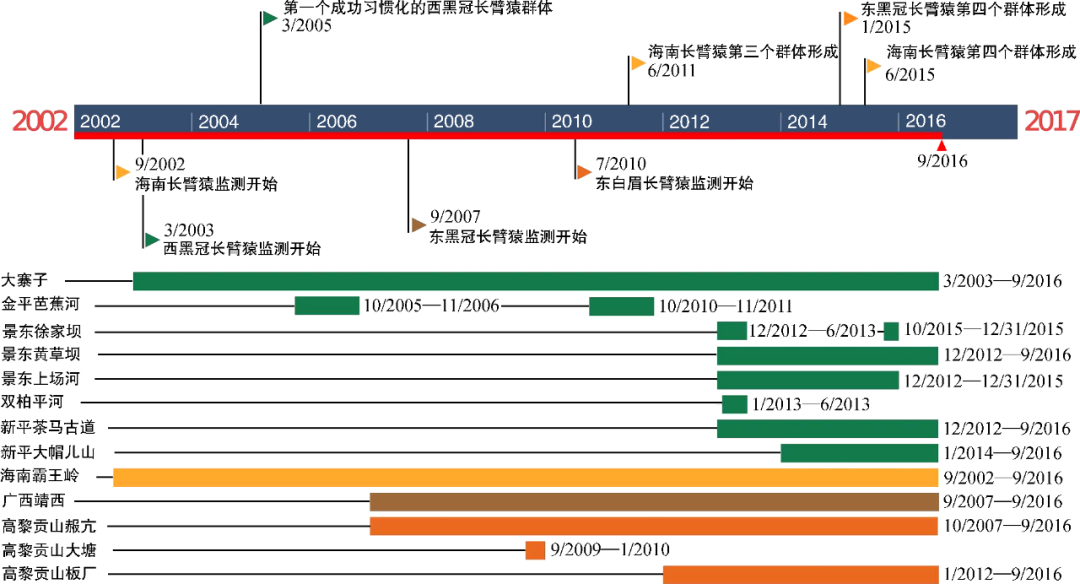
中国长臂猿监测工作时间线 引自管振华等 “中国长臂猿科动物种群监测现状分析”
步入21世纪,我国的科研团队对长臂猿开展了系统而持久的监测和研究,成绩斐然。中国科学院昆明动物研究所蒋学龙研究员团队、中山大学范朋飞教授团队、贵州师范大学周江教授团队,以及西南林业大学、大理大学等科研究机构都在长臂猿的研究和保护中贡献了力量。管振华、黄蓓、马长勇等一批青年才俊陆续在学术界和科普界崭露头角,尤其是在诸多主流媒体、新媒体中的露面让公众有机会了解中国长臂猿的状况。在这个世界长臂猿日,请允许我向上述科研团队表示感谢,并致以崇高的敬意!
参考文献:
邓怀庆,周江. 2015. 海南长臂猿研究现状. 四川动物,34( 4) : 635-640
范朋飞, 黄蓓, 管政华, 蒋学龙. 2010. 西黑冠长臂猿雄性取代前后鸣叫行为的变化. 兽类学报, 30(2): 139-143
范朋飞. 2012. 中国长臂猿科动物的分类和保护现状. 兽类学报,32( 3) : 248-258.
费汉榄, 范朋飞, 向左甫, 马长勇, 张文, 黄涛. 2010. 东黑冠长臂猿鸣叫特征及气象因子对鸣叫的影响. 兽类学报, 30 (4): 377–383.
高罗佩.2015.长臂猿考.上海世纪出版集团中西书局
管振华,阎璐,黄蓓. 2017. 中国长臂猿科动物种群监测现状分析. 四川动物,36(2) : 232 - 238
李兴康,钟旭凯,韦绍干,崔亮伟,范朋飞,管振华,马长勇.2021.群体大小和觅食环境变化对东黑冠长臂猿日移动距离的影响.兽类学报,41 ( 4) : 388-397
李旭,吴建普,周伟,等. 高黎贡山赧亢东白眉长臂猿春秋季食谱及食物结构. 西南林业大学学报,2015,35 ( 2 ) :84-89刘振河,余斯绵,袁喜才. 1984. 海南长臂猿的资源现状. 野生动物,6: 1-4.
马长勇,费汉榄,黄涛,崔亮伟,范朋飞. 2014. 邦亮东黑冠长臂猿日食性与活动节律的季节性变化. 兽类学报,34 ( 2 ) :105 - 114.
马世来,王应祥. 1986. 中国南部长臂猿的分类和分布——附三个新亚种的描记. 动物学研究,7 (4) : 393 - 410.
倪庆永,蒋学龙,王孝伟,等. 西黑冠长臂猿隔离小种群栖息地植被特征与其食性及生境利用. 兽类学报,2015,35 (2) : 119 - 129.
孙国政,倪庆永,黄蓓,等. 2012. 西黑冠长臂猿的种群数量、分布与现状. 林业建设,1: 38-44.
韦绍干,马长勇, 谭武靖, 杨江, 崔亮伟, 范朋飞. 2017. 广西邦亮东黑冠长臂猿新群体的发现及种群数量现状. 兽类学报,37 (3): 233 - 240
吴建普,周伟,李明. 2016. 云南高黎贡山赧亢东白眉长臂猿的鸣叫及其行为. 西部林业科学, 45(2):129-134
周江,陈辈乐,魏辅文. 2008. 海南长臂猿的家族群相遇行为观察. 动物学研究,29( 6) : 667-673.
Fan PF,Huo S. 2009. The northern white-cheeked gibbon ( Nomascus leucogenys) is on the edge of extinction in China. Gibbon Journal,5: 44-52.
Fan PF,Jiang XL. 2009. Predation on giant flying squirrels ( Petaurista philippensis) by black crested gibbons ( Nomascus concolor jingdongensis) at Mt. Wuliang,Yunnan,China. Primates,50(1) : 45-49.
Fan P F,Bartlett T Q,Fei H L,Ma C Y,Zhang W. 2015. Understanding stable bi-female grouping in gibbons: feeding competition and reproductive success. Frontiers in Zoology,12 (1): 5.
Fan, P. F., He, K., Chen, X., Ortiz, A., Zhang, B., et al . 2017 . Description of a new species of Hoolock gibbon (Primates: Hylobatidae) based on integrative taxonomy. American Journal of Primatology, 79(5).
Liu ZH,Zhang YZ,Jiang HS,et al. 1989. Population structure of Hylobates concolor in Bawanglin Nature Reserve,Hainan,China [J]. American Journal of Primatology,19( 4) : 247-254.
Mittermeier R A,Rylands A B,Wilson D E. 2013. Handbook of the Mammals of the World,Vol. 3:Primates. Lynx Edicions,Barcelona.
Thinh V N,Mootnick A R,Thanh V N,Nadler T,Roos C. 2010.A new species of crested gibbon,from the central Annamite Mountain range. Vietnamese Journal of Primatology,4: 1-12.
Trivedi M,Manu S, Balakrishnan S, Biswas J, Asharaf N V K,Umapathy G. 2021. Understanding the phylogenetics of Indian Hoolock Gibbons: Hoolock hoolock and H. leuconedys. InternationalJournal of Primatology,42: 463-477.
Yanqing Guo, Jiang Chang, Ling Han1, Tao Liu1, Gang Li, Paul A. Garber, Ning Xiao, Jiang Zhou. 2020. The Genetic Status of the Critically Endangered Hainan Gibbon (Nomascus hainanus): A Species Moving Toward Extinction. Frontiers in Genetics , 11: 1-11
Yang D H,Zhang J Y,Li C. 1987. Preliminary survey on the population and distribution of gibbons in Yunnan Province. Primates,28(4) : 547 - 549.
来源:
国家动物博物馆
微信号:cwcawx
中国野生动物保护协会
投稿信箱:cwcaweixin@sina.com


阅读
14270

保护地热搜
- 《中国自然资源报》理论版刊发邓侃文章:做好固碳减碳的林业文章 | 阅141194
- 《中国林业》杂志刊发邓侃文章:解读“森林是钱库” | 阅121465
- 物种通用数据 | 阅20620
- 西溪国家湿地公园模式的实践与探索 | 阅17548
- 唐雪琼:后新冠疫情期间的云南自然保护地社区生态旅游发展 | 阅14466
- 我国湿地现状如何?如何解读第25届世界湿地日主题? | 阅14007
- 日本国家公园保护管理观察 | 阅11324
- 红树林该如何保护才科学 | 阅11153
- 2023年生物科技趋势:合成生物占据“C位” | 阅10852
- “两山”理念提出15周年 | 南山国家公园:探索南方丘陵山地自然保护新模式 | 阅9786
| 我也说两句 |
| 版权声明: 1.依据《服务条款》,本网页发布的原创作品,版权归发布者(即注册用户)所有;本网页发布的转载作品,由发布者按照互联网精神进行分享,遵守相关法律法规,无商业获利行为,无版权纠纷。 2.本网页是第三方信息存储空间,阿酷公司是网络服务提供者,服务对象为注册用户。该项服务免费,阿酷公司不向注册用户收取任何费用。 名称:阿酷(北京)科技发展有限公司 联系人:李女士,QQ468780427 网络地址:www.arkoo.com 3.本网页参与各方的所有行为,完全遵守《信息网络传播权保护条例》。如有侵权行为,请权利人通知阿酷公司,阿酷公司将根据本条例第二十二条规定删除侵权作品。 |